

Fizyczne podstawy dekompresji
Teoria dekompresji
Stanisław Bednarek
I. Wstęp
Nam nurkom trudno jest się z tym pogodzić, ale człowiek jest ssakiem typowo lądowym i jego organizm nie jest przystosowany do nurkowania. Na nurka zanurzającego się pod wodę czyha szereg niebezpieczeństw z których najgroźniejsze tkwią w jego organizmie. Dlatego zrozumienie zjawisk zachodzących w jego organizmie i otoczeniu jest warunkiem bezpiecznego nurkowania. Jednym z zagadnień z pogranicza fizyki i fizjologii, którego znajomość jest każdemu nurkowi niezbędna jest zjawisko dekompresji.
Nurek znajdujący się pod wodą poddany jest ciśnieniu otoczenia zwiększonemu o ciśnienie hydrostatyczne znajdującej się ponad nim wody. Zależy więc ono od głębokości nurkowania i wzrasta o 1 at na każde dziesięć metrów głębokości. Podczas typowych nurkowań ciśnienie otoczenia kilkakrotnie przewyższa ciśnienie atmosferyczne. W warunkach zwiększonego ciśnienia następuje zwiększone rozpuszczanie się gazu mieszaniny oddechowej (np. powietrza) we krwi, a za jej pośrednictwem we wszystkich tkankach organizmu. Podczas wynurzania ciśnienie otoczenia, a zarazem mieszaniny oddechowej maleje, dochodzi do sytuacji przesycenia tkanek rozpuszczonym w nich gazem. Największy problem stwarza nam obojętny składnik mieszaniny oddechowej (w powietrzu jest nim azot). Tlen i dwutlenek węgla nie odgrywają w tym procesie większej roli, ze względu na możliwość szybkiego ich transportu przez hemoglobinę (procesy chemiczne), natomiast proces rozpuszczania w tkankach czynnika obojętnego jest procesem czysto fizycznym. Przy nadmiernym przesyceniu tkanki gazem może nastąpić jego wydzielanie w postaci pęcherzyków, które powodują uszkodzenie tkanki w której powstały. Prowadzi to do tzw. choroby dekompresyjnej, dawniej nazywanej kesonową. Jeżeli liczne pęcherzyki gazu przedostaną się do krwiobiegu zamykają światło naczyń włosowatych powodując niedotlenienie organów centralnych lub mięśnia sercowego. Powoduje to tzw. ostrą postać choroby ciśnieniowej. Zanim poznano jej mechanizm, wielu nurków swoją podwodną działalność przypłaciło zdrowiem lub nawet życiem. W celu uniknięcia choroby dekompresyjnej przestrzega się odpowiednio wolnego wynurzania, umożliwiającego wydalenie z organizmu obojętnego czynnika oddechowego przez płuca bez narażania się na wydzielanie w tkankach jego pęcherzyków mających tendencje wzrostowe. Przepis wynurzania musi być odpowiednio prosty, możliwy do kontroli przez człowieka o poważnie ograniczonej przez czynniki zewnętrzne zdolności spostrzegania i rozumowania. W powszechnym użyciu są tzw. tabele dekompresyjne, których skróconą wersję każdy nurek zabiera ze sobą pod wodę. Tabele przewidują tzw. dekompresję stopniowaną. Nurek wynurza się z pod wody z ustaloną prędkością (my przyjmiemy 10m/min) zatrzymując się na kolejnych przystankach dekompresyjnych na określonych głębokościach. Czas pobytu na kolejnych przystankach dekompresyjnych zależy od głębokości i czasu nurkowania, a jego wartości nurek odczytuje z tabeli.
Celem niniejszego opracowania jest przedstawienie prostego modelu umożliwiającego ilościowy opis zjawisk fizycznych zachodzących w tkankach podczas nurkowania. Został on zaproponowany przez grupę prof. Bűhlmana z Zűrichu w roku 1986. Model jest nadal aktualny, służy do wyliczania tabel dekompresyjnych, które są w powszechnie używane przez nurków sportowych w Europie. Podobne, choć nieco uproszczone procedury numeryczne są zaimplementowane w większości komputerów nurkowych, które pojawiły się na światowym rynku w latach 90-tych.
II. Podstawowe prawa fizyczne
Przedstawiona poniżej teoria dekompresji bazuje na kilku prawach fizycznych, których znajomość jest niezbędna dla zrozumienia zagadnienia. Ponieważ ich zastosowanie jest dość nietypowe, pozwolę sobie na przypomnienie ich.
1.Prawo ciśnień parcjalnych (Daltona)
Aktywność chemiczna, oraz niektóre własności fizyczne np. rozpuszczalność w cieczach poszczególnych składników mieszaniny gazowej zależy od jego ciśnienia parcjalnego Pi , które jest iloczynem procentowego udziału interesującego nas składnika Ci przez ciśnienie całkowite mieszaniny P: Pi=CiP (1)
Na przykład ciśnienie parcjalne azotu w powietrzu atmosferycznym na poziomie morza PN=78%*1at = 0.78at. Naturalnie suma ciśnień parcjalnych wszystkich składników mieszaniny jest równa ciśnieniu całkowitemu. 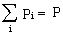 (2)
(2)
2. Prawo rozpuszczalności gazu w cieczach (Henry'ego).
Maksymalna ilość składnika mieszaniny gazowej, która może rozpuścić się w cieczy po dojściu do stanu równowagi (nasycająca ciecz) jest proporcjonalna do ciśnienia parcjalnego tego składnika w fazie gazowej znajdującej się nad powierzchnią cieczy. Dzięki temu ilość gazu rozpuszczonego w cieczy określać można podając ciśnienie nasycenia, czyli ciśnienia przy jakim ciecz byłaby z gazem w równowadze.
3. Rozpuszczanie się gazu w dwóch nie mieszających się cieczach.
Jeżeli dwie nie mieszające się ciecze (poszczególne tkanki człowieka traktowane są w tym modelu jako ciecze) są w równowadze z fazą gazową, wówczas stężenia gazu rozpuszczone go w nich są proporcjonalne do ciśnienia, ale nie są równe. Współczynniki proporcjonalności, różne dla różnych cieczy (tkanek) noszą nazwę współczynników podziału Kj. Cj=KjP (3)
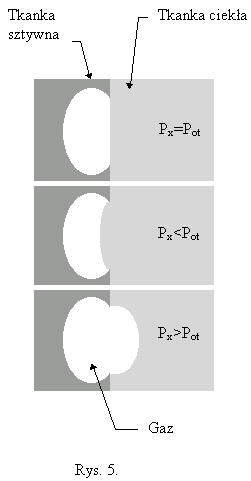
III. Równanie nasycania (odsycania)
Połączenie praw 2. i 3. pozwala na ułożenie równania opisującego proces nasycania (odsycania) tkanek podczas nurkowania. Przedyskutujmy procesy zachodzące w tkance posługując się modelem przedstawionym na rysunku poniżej.
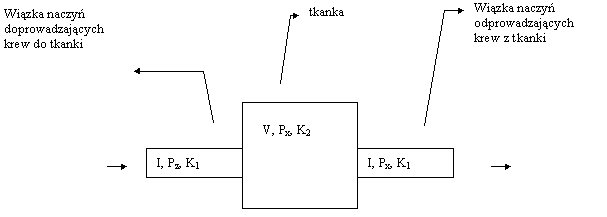
Załóżmy, że tkanka ma objętość V,a aktualne ciśnienie nasycenia tkanki gazem oznaczmy przez Px. Do tkanki dopływa krew nasycona w płucach do ciśnienia otoczenia Pot pomniejszonego przez ciśnienie parcjalne pary wodnej w pęcherzykach płucnych Pw=0.063at. Jeżeli jako zawartość gazów obojętnych w powietrzu (azot+argon) przyjmijmy C=0.79. Wówczas do tkanki dopływa krew o ciśnieniu nasycenia: Pz=(Pot - Pw)C (4)
Niech w jednostce czasu wpływa do niej objętość I krwi. Z tkanki wypływa krew z prędkością również I, ale ze względu na dużą powierzchnię naczyń włosowatych będących z nią w kontakcie, ciśnienie nasycenia krwi wypływającej jest równe Px. Przyjmijmy współczynniki podziału dla krwi i tkanki odpowiednio K1 i K2. Bilansując ilość dopływającego, odpływającego i pozostawianego w tkance gazu w ciągu czasu dt, otrzymujemy:
I K1Pzdt = VK2dPx + IK1Pxdt (5) a stąd po podzieleniu obu stron równania przez dt i przekształceniu, otrzymujemy równanie różniczkowe opisujące zależność czasową ciśnienia nasycenia tkanki.
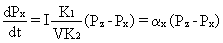 (6)
(6)
Współczynnik ax stanowi wypadkową objętości tkanki, perfuzji - czyli szybkości przepływu krwi przez tkankę, oraz współczynników podziału K1 dla krwi i K2 dla tkanki. Jest charakterystyczny dla danej tkanki, a jego sens fizyczny możemy wydobyć rozwiązując równanie (5) dla schodkowego profilu nurkowania. Zakładamy że ciśnienie nasycenia tkanki w chwili t=0 wynosi P0 i zwiększamy ciśnienie otoczenia tak, by ciśnienie parcjalne gazu we krwi wyniosło Pz. Wówczas
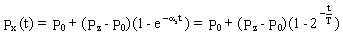 (7)
(7)
Proces nasycania (odsycania) przebiega wykładniczo. T=ln2/ax nosi nazwę okresu półnasycenia i jest czasem po którym ciśnienie nasycenia tkanki osiąga połowę wartości pomiędzy ciśnieniem początkowym a aktualnie działającym. Okres półnasycenia jest oczywiście różny dla różnych tkanek. Różnice mogą wystąpić w ramach tej samej tkanki w miejscach różnie ukrwionych. Przebieg ciśnienia nasycenia podczas nurkowania dla kilku tkanek o różnym okresie półnasycenia jest przedstawiony na Rysunku 2. Dla lepszej ilustracji założono profil nurkowania polegający na gwałtownym zanurzeniu na głębokość 40m i pobyt na stałe j głębokości. Sposób identyfikacji okresu półnasycenia zilustrowano liniami przerywanymi.
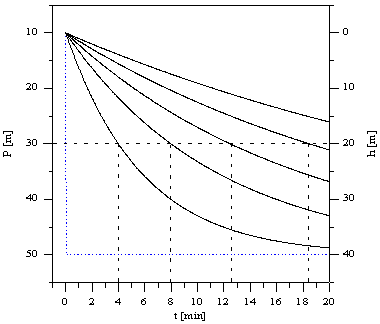 Rys. 2.Krzywe nasycenia tkanek w funkcji czasu. Krzywe ciągłe określają przebiegi ciśnienia nasycenia kilku wybranych tkanek. Linia kropkowana oznacza profil nurkowania. Na prawej skali ododczytujemy głębokość nurkowania, na lewej odpowiadające jej ciśnienie. Jako jednostkę ciśnienia przyjęto metr słupa wody.
Rys. 2.Krzywe nasycenia tkanek w funkcji czasu. Krzywe ciągłe określają przebiegi ciśnienia nasycenia kilku wybranych tkanek. Linia kropkowana oznacza profil nurkowania. Na prawej skali ododczytujemy głębokość nurkowania, na lewej odpowiadające jej ciśnienie. Jako jednostkę ciśnienia przyjęto metr słupa wody.
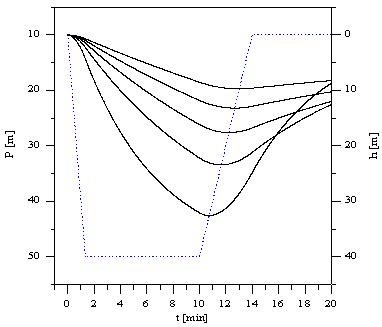
Rys. 3.Krzywe nasycania i odsycania tkanek podczas trapezoidalnego profilu nurkowania. Przyjęto prędkość zanurzania 30m/min, wynurzania 10m/min. Przyjęto oznaczenia zgodne z Rys. 2.
Na wykresie przedstawiono ciśnienia całkowite mieszaniny oddechowej, łącznie z tlenem i azotem. Jako jednostkę ciśnienia przyjęto metr słupa wody i odwrócono skalę. Ciśnienie wzrasta w dół wykresu. Umożliwia to porównanie aktualnych ciśnień nasycenia z ciśnieniem otoczenia panującym na głębokości na jakiej znajduje się nurek. Zauważmy, że na początku i nurkowania kiedy nurek znajdował się na powierzchni ciśnienie otoczenia było równe 10 m słupa wody. jest to ciśnienie atmosferyczne. Na Rysunku 3 przedstawione zostały przebiegi nasycania tkanek podczas zanurzania i pobytu na maksymalnej głębokości i odsycania podczas wynurzania. Przyjęto trapezoidalny profil nurkowania. Zanurzenie i wynurzenie odbywa się ze skończoną prędkością. Założono prędkość wynurzania 10m na minutę. Prędkość zanurzania przyjęto większą. Zauważmy, że tkanki "szybkie" , czyli tkanki o małym okresie półnasycenia nasycają się szybciej i osiągają większe maksymalne ciśnienia nasycenia, ale odsycają się również szybciej i przy wynurzaniu ich ciśnienie nasycenia staje się mniejsze od ciśnień nasycenia tkanek "wolniejszych". Tkanki szybkie "śledzą" profil nurkowania. Przy odpowiednio małej prędkości wynurzania ok. 10 m/min tkanki szybkie nie sprawiają żadnych kłopotów, zawarty w nich gaz usuwany jest za pośrednictwem krwioobiegu przez płuca. Warunkiem bezpiecznego wynurzenia jest usunięcie tą drogą gazu z wszystkich tkanek organizmu. Przeanalizujmy problem alternatywnej, niekorzystnej dla organizmu możliwości wydzielania gazu z tkanek, mianowicie mechanizm powstawania i wzrostu pęcherzyków gazowych.
IV. Warunki wzrostu pęcherzyków gazowych
Wyobraźmy sobie pęcherzyk gazu w cieczy, w postaci kuli o promieniu r. Wewnątrz pęcherzyka panuje ciśnienie równe ciśnieniu nasycenia cieczy, z którą gaz zawarty w pęcherzyku musi być w równowadze. Z zewnątrz na pęcherzyk działa ciśnienie otoczenia powiększone o ciśnienie związane z napięciem powierzchniowym na granicy gaz - ciecz. To ostatnie zależy od promienia pęcherzyka i współczynnika napięcia powierzchniowego s. Dane jest wyrażeniem:
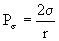 (8)
(8)
W połączeniu z ciśnieniem otoczenia daje ono ciśnienie zewnętrzne działające na pęcherzyk gazu: Pz=Pot+ Ps (9)
Jeżeli ciśnienie zewnętrzne Pz jest większe od ciśnienia nasycenia tkanki Px, pęcherzyk gazu będzie zmniejszał swoją objętość i jeżeli powstał to zniknie. W przeciwnym przypadku jego objętość rośnie. Warunki równowagi zilustrowane są na Rysunku 4.
Rys. 4.
Px
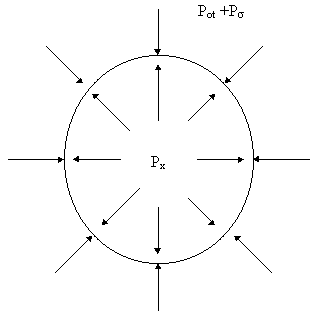
Dzięki napięciu powierzchniowemu ciecz może być przesycona, czyli ciśnienie nasycenia może przekraczać ciśnienie zewnętrzne, a mimo to pęcherzyki gazu nie powstają. Różnicę ciśnienia nasycenia i ciśnienia otoczenia nazywamy ciśnieniem przesycenia: D Px=Px-Pot (10)
W idealnie czystej i jednorodnej cieczy pęcherzyki gazowe nie powinny powstać, ponieważ Ps dąży do Ydla r bliskiego 0 i D Px nie przekracza Ps dla nieskończenie małego promienia pęcherzyka.
Jednakże w tkankach istnieją tzw. zarodzie, powodujące samoistne pojawianie się pęcherzyków gazu o pewnym promieniu r0. Istnieje więc pewne przesycenie krytyczne: D Px=Px-Pot=2s /r0, (11)
po przekroczeniu którego w tkance następuje produkcja pęcherzyków mających tendencje wzrostowe. To przesycenie krytyczne jest różne dla różnych tkanek ze względu na różne napięcie powierzchniowe oraz różny charakter zarodzi.
V. Prosty model zarodzi
Zarodzie będące przyczyną pojawiania się w cieczy przesyconej pęcherzyków gazu mogą mieć różnorodny charakter. Aby problem zarodzi uczynić mniej tajemniczym, przeanalizujmy model tzw. "kieszeni gazowej", który określa jeden z możliwych mechanizmów ich powstawania. Jeżeli na brzegu naczynia krwionośnego znajduje się zagłębienie wypełnione gazem, wówczas w warunkach równowagi ciśnienia nasycenia krwi z ciśnieniem otoczenia, granica ciecz-powietrze tworzy płaską powierzchnie zamykająca zagłębienie. Przy wzroście ciśnienia otoczenia powierzchnia ta uwypukla się w kierunku wgłębienia. Jednocześnie na skutek działania napięcia powierzchniowego zawartość gazu w "kieszeni rośnie dążąc do zmniejszenia powierzchni kontaktu gaz-ciecz. Podczas zmniejszania ciśnienia otoczenia gaz rozpręża się i w warunkach przesycenia roztworu kieszeń może być źródłem znacznej ilości kolejno odrywających się pęcherzyków gazu, o pewnym minimalnym promieniu zależącym od promienia
"szyjki kieszeni".
VI. Matematyczny model organizmu (Bűhlmana)
Złożoność procesów zachodzących w organizmie nurka podczas zanurzania i wynurzania nie pozwala na pełny ich teoretyczny opis. Przeprowadzona powyżej dyskusja nie jest pełną teorią, może być traktowana jedynie jako ich przybliżoną ilustrację. Jej celem było wprowadzenie i wytłumaczenie podstawowych pojęć i terminów, które użyte zostały w zaproponowanym przez Bűhlmana modelu matematycznym.
Zanim przejdziemy do konkretyzacji modelu wprowadzę jeszcze jedno pojęcie, które będzie w nim wykorzystane. Przez Ptol oznaczmy tolerowalne przez rozpatrywaną tkankę ciśnienie otoczenia, czyli minimalne ciśnienie zewnętrzne przy jakim nie powstają w niej jeszcze pęcherzyki powietrza. Określa ono do jakiej minimalnej głębokości może wynurzyć się nurek przy aktualnym stanie nasycenia jego tkanki. Gdybyśmy znali przesycenia krytyczne, ciśnienie to dane by było wyrażeniem:
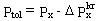
W modelu matematycznym Buhlman ograniczył liczbę występujących w organizmie, T półnasycenia okresy im odpowiadające oraz j, numery Ich 16. do tkanek parametrami nas interesującymi się różniących>sub>j zestawione są w pierwszej i drugiej kolumnie Tabeli I. W trzeciej i czwartej kolumnie znajdują się odpowiadające tkankom współczynniki aj,
i bj przy pomocy których sparametryzowane zostało tolerowalne przez daną tkankę ciśnienie otoczenia Pjtol, którego sens zawarty jest w równaniu (7), ale liczone jest przy pomocy wyrażenia:
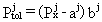 | (13) |
Parametry Tj, aj, i bj są parametrami dopasowania modelu i wyznaczone zostały na podstawie materiału doświadczalnego zgromadzonego przez autora modelu (Bűhlman). Z punktu widzenia fizyki model ten ma zbyt wiele parametrów dopasowania. Jednakże ponieważ opisywany układ jakim jest organizm ludzki nie jest możliwe jego uproszczenie i musi być zaakceptowany.
Użyte w modelu parametry
charakteryzujące tkanki.
| j | Tj min | aj at | bj |
| 1 | 4.0 | 1.906 | 0.810 |
| 2 | 8.0 | 1.450 | 0.810 |
| 3 | 12.5 | 1.040 | 0.810 |
| 4 | 18.5 | 0.880 | 0.830 |
| 5 | 27.0 | 0.717 | 0.845 |
| 6 | 38.3 | 0.577 | 0.860 |
| 7 | 54.3 | 0.468 | 0.870 |
| 8 | 77.0 | 0.441 | 0.903 |
| 9 | 109.0 | 0.415 | 0.908 |
| 10 | 146.0 | 0.416 | 0.939 |
| 11 | 187.0 | 0.369 | 0.946 |
| 12 | 239.0 | 0.369 | 0.946 |
| 13 | 305.0 | 0.255 | 0.962 |
| 14 | 390.0 | 0.255 | 0.962 |
| 15 | 498.0 | 0.255 | 0.962 |
| 16 | 635.0 | 0.255 | 0.962 |
VII. Metoda liczenia tabel dekompresyjnych i sposób posługiwania się nimi
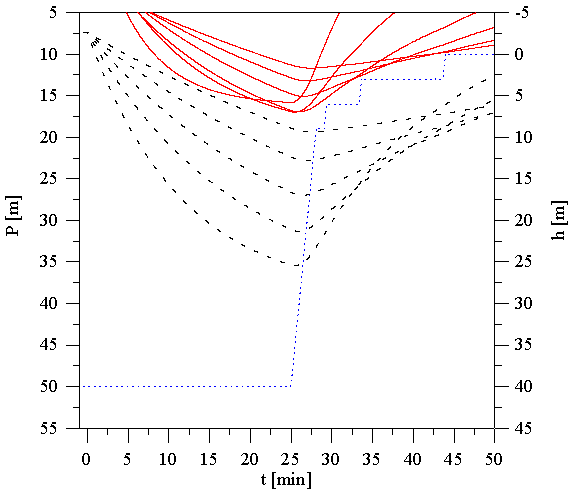
Rysunek 6 przedstawia przykładowy przebieg trwającego 25 min nurkowania do głębokości 40 m z poprawnie zaplanowanym wynurzeniem. Rysunek 7 jest powiększonym fragmentem zawierającym tzw. przystanki dekompresyjne. Przy obliczaniu tabel dekompresyjnych zakładamy trapezoidalny profil nurkowania, niezależnie od jego faktycznego przebiegu. Zakładamy, że nurek zanurza się od razu na maksymalną osiągnięta podczas całego nurkowania głębokość i przebywa tam do momentu rozpoczęcia wynurzenia. Następnie wynurza się z prędkością 10m na min. Profil nurkowania przedstawia krzywa kropkowana. Krzywe przerywane określają znane nam z poprzednich rysunków ciśnienia nasycenia pięciu "najszybszych tkanek". Leżące powyżej krzywe ciągłe przedstawiają odpowiadające im ciśnienia tolerowalne. Wynurzenie powinno być tak zaplanowane, aby ciśnienie otoczenia na głębokości na której znajduje się nurek nie było mniejsze od ciśnień tolerowanych przez wszystkie tkanki. W przypadku gdy ciśnienie otoczenia osiągnie wartość najniższego ciśnienia tolerowalnego, nurek musi się zatrzymać na tzw. przystanku dekompresyjnym i odczekać, aż ciśnienia tolerowalne obniżą się i pozwolą mu wynurzyć się do głębokości odpowiadającej następnemu przystankowi dekompresyjnemu. Głębokości przystanków dekompresyjnych standardowo są wielokrotnościami 3 m, czasy postoju na przystankach są przyjęto jako wielokrotności 1 min.
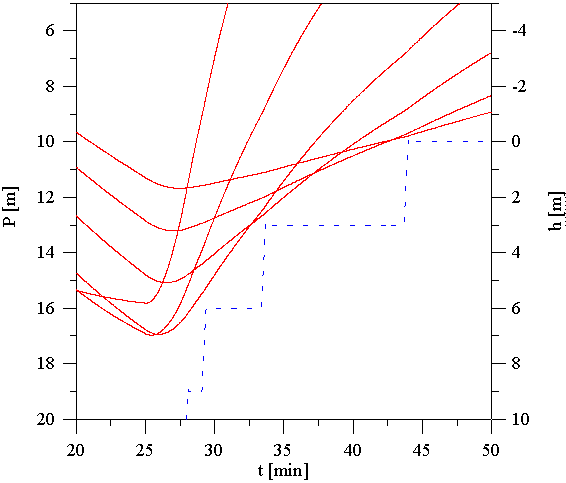
Tkanka, której ciśnienie tolerowane spowodowało zatrzymanie nurka na przystanku dekompresyjnym nosi nazwę tkanki kontrolnej. W różnych fazach nurkowania tkanki kontrolne się zmieniają. Spójrzmy na Rysunek 6. Pierwszy przystanek dekompresyjny wystąpił na głębokości 9m. Do zatrzymania nurka na nim zmusiła go możliwość przekroczenia ciśnienia tolerowanego przez tkankę o okresie półnasycenia 12.5 min, dla tego przystanku kontrolną była tkanka ?18.5-minutowa?. Również na przystanku na głębokości 6m, kontrolną była ta sama tkanka. Na przystanku na głębokości 3m kontrolnymi były tkanki 12.5-min i 18.5-min, a przy wynurzaniu na powierzchnię tkanki 27-min i 38.3-min. Początkowe fazy wynurzania kontrolowane są przez ?szybkie? tkanki, końcowe przez tkanki powolne.
VIII. Uwagi końcowe
1.Jeżeli zsumujemy ze sobą czasy postoju na poszczególnych przystankach dekompresyjnych i dodamy do nich czas wypływania na powierzchnię z prędkością ograniczoną do 10 m/min, okaże się, że łączny czas bezpiecznego wynurzania wyniósł 18 min. Był więc porównywalny z czasem samego nurkowania, który założyliśmy 25 min. Przy nurkowaniach głębszych, lub dłużej trwających wymuszony koniecznością dekompresji czas wynurzania wielokrotnie przekracza czas nurkowania. Ma to szczególne znaczenie przy planowaniu nurkowania, gdyż nurek może zabrać pod wodę tylko skończony zapas powietrza i nie może mu go zabraknąć na poprawne i zgodne z zasadami dekompresji wynurzenie.
2. Po zakończeniu nurkowania nie wszystkie tkanki są odsycone. Zanurzając się ponownie musimy mieć świadomość, że "powolne" tkanki startują z początkowym ciśnieniem nasycenia większym niż przy pierwszym nurkowaniu. W tym wypadku musimy więc posługiwać się tabelami wielokrotnego nurkowania. Tabelami opracowanymi dla pierwszego nurkowania możemy się posłużyć jedynie w przypadku około 24 godzinnej przerwy pomiędzy nurkowaniami.
3. Ze względu na przesycenie "powolnych" tkanek nie możemy bezpośrednio po nurkowaniu podróżować samolotem, gdyż na typowych wysokościach lotu ciśnienie otoczenia jest znacznie niższe niż na poziomie morza. Tabele dekompresyjne naogół określają czas po którym można takie podróże odbywać.
4. Również nurkowanie w jeziorach górskich wymaga stosowania innych tabel dekompresyjnych, ze względu na niższe ciśnienie atmosferyczne na powierzchni.
5. Tabele dekompresyjne przewidują zazwyczaj znacznie dłuższy czas dekompresji niż faktycznie jest niezbędny, ze względu na założony trapezoidalny profil nurkowania zakładający pobyt nurka na maksymalnej głębokości przez cały czas nurkowania. Podczas typowych nurkowań na maksymalnej głębokości nurek przebywa znacznie krócej. Zanurza się ze skończoną prędkością, zatrzymuje się podczas zanurzania lub wraca na mniejszą głębokość. Uwzględnienie prawdziwych profili nurkowania nie jest możliwe w postaci tabel. W tym celu nurek zabiera ze sobą pod wodę komputer nurkowy mierzący czas i głębokość zanurzenia i na bieżąco zliczający ciśnienia nasycenia poszczególnych tkanek. Użycie komputera nurkowego optymalizuje czas dekompresji i jest znacznie mniej uciążliwe od posługiwania się tabelami dekompresyjnymi.
Jak zapisać się na kurs nurkowania!
Kursy nurkowania prowadzimy w Krakowie. Zależy Ci na szybkim terminie - skontaktuj się z instruktorem telefonicznie lub e-mail. Zawsze możesz zadzwonić i zapytać. Preferujemy kontakt telefoniczny. Więcej.